ARTÍCULO
sábado, 30 de julio de 2011
Video del artículo Radicales libres en la Biomedicina y Estrés Oxidativo
sábado, 23 de julio de 2011
VISIÓN
VISIÓN
La luz visible es una de las formas como se desplaza la energía. Las ondas de luz son el resultado de vibraciones de campos eléctricos y magnéticos, y es por esto que son una forma de radiación electromagnética (EM). La luz visible es tan sólo uno de los muchos tipos de radiación EM, y ocupa un pequeño rango de la totalidad del espectro electromagnético . 2

{kind=link}
El ojo en su conjunto, llamado globo ocular, es una estructura esférica de aproximadamente 2,5 cm de diámetro con un marcado abombamiento sobre su superficie delantera. La parte exterior, o la cubierta, se compone de tres capas de tejido: la capa más externa o esclerótica tiene una función protectora, cubre unos cinco sextos de la superficie ocular y se prolonga en la parte anterior con la córnea transparente; la capa media o úvea tiene a su vez tres partes diferenciadas: la coroides —muy vascularizada, reviste las tres quintas partes posteriores del globo ocular— continúa con el cuerpo ciliar, formado por los procesos ciliares, y a continuación el iris, que se extiende por la parte frontal del ojo. La capa más interna es la retina, sensible a la luz. 1
La córnea es una membrana resistente, compuesta por cinco capas, a través de la cual la luz penetra en el interior del ojo. Por detrás, hay una cámara llena de un fluido claro y húmedo (el humor acuoso) que separa la córnea de la lente del cristalino. En sí misma, la lente es una esfera aplanada constituida por un gran número de fibras transparentes dispuestas en capas. Está conectada con el músculo ciliar, que tiene forma de anillo y la rodea mediante unos ligamentos.1
El músculo ciliar y los tejidos circundantes forman el cuerpo ciliar y esta estructura aplana o redondea la lente, cambiando su longitud focal. El iris es una estructura pigmentada suspendida entre la córnea y el cristalino y tiene una abertura circular en el centro, la pupila. El tamaño de la pupila depende de un músculo que rodea sus bordes, aumentando o disminuyendo cuando se contrae o se relaja, controlando la cantidad de luz que entra en el ojo.1
Por detrás de la lente, el cuerpo principal del ojo está lleno de una sustancia transparente y gelatinosa (el humor vítreo) encerrado en un saco delgado que recibe el nombre de membrana hialoidea. La presión del humor vítreo mantiene distendido el globo ocular. La retina es una capa compleja compuesta sobre todo por células nerviosas. Las células receptoras sensibles a la luz se encuentran en su superficie exterior detrás de una capa de tejido pigmentado. Estas células tienen la forma de conos y bastones y están ordenadas como los fósforos de una caja. Situada detrás de la pupila, la retina tiene una pequeña mancha de color amarillo, llamada mácula lútea; en su centro se encuentra la fóvea central, la zona del ojo con mayor agudeza visual. La capa sensorial de la fóvea se compone sólo de células con forma de conos, mientras que en torno a ella también se encuentran células con forma de bastones. Según nos alejamos del área sensible, las células con forma de cono se vuelven más escasas y en los bordes exteriores de la retina sólo existen las células con forma de bastones. El nervio óptico entra en el globo ocular por debajo y algo inclinado hacia el lado interno de la fóvea central, originando en la retina una pequeña mancha redondeada llamada disco óptico. Esta estructura forma el punto ciego del ojo, ya que carece de células sensibles a la luz. El ojo humano es receptivo a la radiación electromagnética que denominamos luz visible y que notaremos como distribución espectral, siendo la longitud de onda. Al intervalo de valores de que va de 350nm (nanometros) a 780nm lo llamaremos luz visible. La distribución de la sensibilidad del ojo a las distintas longitudes de onda tiene forma de campana con un valor máximo para los conos en torno a los 600 nm y un máximo para los bastones en torno a los 500 nm. 1
Los conos son células sensibles a la luz que se encuentran situadas en la retina de los vertebrados, en la llamada capa de conos y bastones. Reciben este nombre por su forma conoidea. Estas células son las responsables de la visión en colores. En la zona central de la retina (fóvea), la cantidad de conos es mayor, su número desciende a medida que nos acercamos a la perifería. En la especie humana y en muchos otros primates, existen tres tipos diferentes de conos, cada uno de ellos es sensible de forma selectiva a la luz de una longitud de onda determinada, verde, roja y azul. Esta sensibilidad especifica se debe a la presencia de unas sustancias llamadas opsinas. La eritropsina tiene mayor sensibilidad para las longitudes de onda largas de alrededor de 560 nanómetros (luz roja), la cloropsina para longitudes de onda medias de unos 530 nanómetros (luz verde) y por último la cianopsina con mayor sensibilidad para las longitudes de onda pequeñas de unos 430 nanómetros (luz azul). El cerebro interpreta los colores a partir de la razón de estimulación de los tres tipos de conos. 1
Existen mamíferos nocturnos que poseen solamente uno de estos pigmentos, mientras que algunas aves y reptiles tienen cuatro y son capaces de detectar la luz ultravioleta no visible para los humanos. Las señales generadas en los conos se transmiten en la retina a las células bipolares que conectan con las células ganglionares de donde parte el nervio óptico que envia la informacion al cerebro.Los bastones son células fotorreceptoras de la retina responsables de la visión en condiciones de baja luminosidad. Presentan una elevada sensibilidad a la luz aunque se saturan en condiciones de mucha luz y no detectan los colores. Se ubican en casi toda la retina exceptuando la fóvea. Contienen rodopsina, que es una proteína que presenta una mayor sensibilidad a las longitudes de onda cercanas a 500 nanómetros, es decir, a la luz verde azulada. 2
Los conos son células sensibles a la luz que se encuentran situadas en la retina de los vertebrados, en la llamada capa de conos y bastones. Reciben este nombre por su forma conoidea. Estas células son las responsables de la visión en colores. En la zona central de la retina (fóvea), la cantidad de conos es mayor, su número desciende a medida que nos acercamos a la perifería. En la especie humana y en muchos otros primates, existen tres tipos diferentes de conos, cada uno de ellos es sensible de forma selectiva a la luz de una longitud de onda determinada, verde, roja y azul. Esta sensibilidad especifica se debe a la presencia de unas sustancias llamadas opsinas. La eritropsina tiene mayor sensibilidad para las longitudes de onda largas de alrededor de 560 nanómetros (luz roja), la cloropsina para longitudes de onda medias de unos 530 nanómetros (luz verde) y por último la cianopsina con mayor sensibilidad para las longitudes de onda pequeñas de unos 430 nanómetros (luz azul). El cerebro interpreta los colores a partir de la razón de estimulación de los tres tipos de conos. 1
Existen mamíferos nocturnos que poseen solamente uno de estos pigmentos, mientras que algunas aves y reptiles tienen cuatro y son capaces de detectar la luz ultravioleta no visible para los humanos. Las señales generadas en los conos se transmiten en la retina a las células bipolares que conectan con las células ganglionares de donde parte el nervio óptico que envia la informacion al cerebro.Los bastones son células fotorreceptoras de la retina responsables de la visión en condiciones de baja luminosidad. Presentan una elevada sensibilidad a la luz aunque se saturan en condiciones de mucha luz y no detectan los colores. Se ubican en casi toda la retina exceptuando la fóvea. Contienen rodopsina, que es una proteína que presenta una mayor sensibilidad a las longitudes de onda cercanas a 500 nanómetros, es decir, a la luz verde azulada. 2
Los bastones se conectan en grupo y responden a los estímulos que alcanzan un área general, pero no tienen capacidad para separar los pequeños detalles de la imagen visual. La diferente localización y estructura de estas células conduce a la división del campo visual del ojo en una pequeña región central de gran agudeza y una zona periférica de menor agudeza, pero con gran sensibilidad a la luz. Así, durante la noche, los objetos se pueden ver por la parte periférica de la retina cuando son invisibles para la fóvea central. Los bastones son más delgados que los conos, el diámetro de sus segmentos internos es de aproximadamente 2 micras. Los segmentos externos de los bastones están formados por discos membranosos aislados de la membrana plasmática, donde se encuentra la rodopsina. Estos discos están continuamente renovándose. Los discos antiguos se van desplazando hacia la zona del epitelio pigmentario, donde son fagocitados y convertidos en fagosomas durante el ciclo diurno, sobre todo al amanecer. Estas células son muy sensibles, capaces de detectar la energía de un sólo fotón y las responsables por tanto de que sea posible la visión nocturna. 1
En los fotorreceptores se produce una transducción foto-quimio-eléctrica que da lugar a que en la terminal sináptica se libere mayor o menor cantidad de NT en relación con la magnitud del potencial receptor. La estimulación de los fotorreceptores se inicia por la absorción de la luz por el pigmento visual y el efecto fotoquímico correspondiente; ello lleva consigo cambios de permeabilidad iónica y génesis de potencial receptor que, desde el segmento externo, pasa al segmento interno y se transmite a la región sináptica donde, mediada por un transmisor, la señal alcanza otras neuronas retinianas (bipolares y horizontales). Los fotorreceptores son distintos al resto de receptores sensoriales pues no detectan impulsos nerviosos típicos; sin embargo, en los bastones y conos, al ser alcanzados por la luz, se establecen unas ciertas condiciones físico-químicas que van a desencadenar el impulso de otras células nerviosas. Donde si se detectan los impulsos nerviosos es en el nervio óptico, por fibras nerviosas de las células ganglionares. Sin lugar a equivocarnos, la fotoquímica de la visión es el mecanismo más complicado y preciso de los sentidos. Este complejo mecanismo lo estudiamos en tres apartados: ciclo del pigmento visual, generación del potencial receptor y adaptación a la luz y oscuridad.Las teorías sobre la visión de los colores deben de explicar los hechos señalados. La más aceptada es la siguiente: Teoría de Young-Helmholtz. En los conos existirían tres sustancias químicas sensibles, una a la luz roja, otra a la verde y la tercera a la violeta. Esta sustancia se descompone y estimula ciertas fibras nerviosas que llevan sus impulsos a la corteza visual. La sensación de rojo, verde o violeta se debería a la estimulación de la sustancia correspondiente por la luz; la de los otros colores del espectro y la del blanco, por la estimulación de los tres receptores en grado variable; la del negro, por la falta de estimulación. Esta teoría explica bien algunos hechos: a) la ceguera a un color, que se debería a la falta de la sustancia correspondiente; b) la mezcla de colores, que se relacionaría con la estimulación de varias de ellas; c) las posimágenes positivas, que se explicarían por la continuación de la descomposición de la sustancia al cesar el estímulo, o las negativas, que derivarían de la disminución de una sustancia y de la persistencia de las otras dos, que se excitarían con los colores restantes al mirar la superficie blanca. Esta teoría tiene también algunas objeciones serias. Por ejemplo, no explica la visión del blanco y el gris en la periferia de la retina ni explica la visión del amarillo más allá de la zona del rojo y el verde, que son sus colores formadores. Finalmente no se comprende la ceguera total al color cuando hay visión conservada de los objetos. 2


Bibliografía
SENTIDO DEL GUSTO
GUSTO
EL GUSTO es el sentido que nos permite saborear las cosas. Se experimenta la sensación del gusto por medio de la lengua. 1
La lengua es un órgano muscular, movible, que además de experimentar la sensación del gusto sirve para otras funciones como el habla, el masticamiento y el tragar de los alimentos. 1

Las células en que ocurren los primeros eventos de la tranduccion gustativa son células neuroepiteliales que en los vertebrados terrestres se encuentran solamente en la cavidad bucal formando parte de los botones gustativos, distribuidos en la superficie del epitelio gustativo. Los botones gustativos se encuentran preferencialmente en las papilas del epitelio lingual. Los vertebrados marinos poseen además receptores gustativos distribuidos en la superficie corporal. 2
En los mamíferos se aprecian tres tipos de papilas gustativas: fungiformes, foliformes y valladas. Las papilas fungiformes contienen entre uno y nueve botones gustativos cada una y están ubicadas en los dos tercios anteriores de la lengua. Las papilas foliformes y valladas, localizadas en la zona posterior y lateral de la lengua respectivamente, despliegan docenas de botones gustativos. 2
Los botones gustativos están compuestos de 30 a 150 células. De acuerdo al criterio clásico, el botón gustativo poseería tres tipos celulares: las células sostén, ubicadas en la periferia, los receptores gustativos, de forma elongada, que tienen una posición central y despliegan microvellosidades en su porción apical, y las células basales que están en constante división y poseen la capacidad de diferenciarse en células gustativas cuya vida media es de diez días a dos semanas. 2

Para que la lengua sea sensible a algún objeto es necesario que éste se encuentre húmedo. Si no lo está, entonces se humedecerá con ayuda de la saliva que secreta la misma lengua. Esta es otra función que tiene la saliva. 2
El objeto ya húmedo se deposita en la lengua y sus moléculas entran en contacto con los microvili, es decir, los extremos de las fibras de las yemas gustativas. De manera análoga a lo que ocurre en el olfato, se lleva a cabo una reacción química que provoca que se desencadene una respuesta de la célula que nos da la sensación del gusto del objeto en cuestión. 2
Hay que mencionar, además del efecto químico producido en las yemas gustativas, que la sensación del gusto queda determinada por otras propiedades del objeto que producen sensaciones táctiles. Estas propiedades son tanto físicas como químicas. Las primeras de dichas propiedades son el tamaño de la partícula, su textura, su consistencia y su temperatura. Las segundas, las propiedades químicas, producen las sensaciones de frialdad de la menta, lo picoso del chile, etcétera.2
La lengua humana no es sensible de manera uniforme a lo largo de su superficie. Así, la parte media no es sensible al gusto como lo son otras partes. En la figura 60 vemos que la punta de la lengua es sensible principalmente a lo dulce. A los lados y en la parte anterior de la lengua se encuentra la sensibilidad a lo salado. La lengua es principalmente sensible a los sabores agrios a los lados, pero en la parte posterior. Finalmente, la parte posterior central de la lengua es sensible principalmente a los sabores amargos.2
El objeto ya húmedo se deposita en la lengua y sus moléculas entran en contacto con los microvili, es decir, los extremos de las fibras de las yemas gustativas. De manera análoga a lo que ocurre en el olfato, se lleva a cabo una reacción química que provoca que se desencadene una respuesta de la célula que nos da la sensación del gusto del objeto en cuestión. 2
Hay que mencionar, además del efecto químico producido en las yemas gustativas, que la sensación del gusto queda determinada por otras propiedades del objeto que producen sensaciones táctiles. Estas propiedades son tanto físicas como químicas. Las primeras de dichas propiedades son el tamaño de la partícula, su textura, su consistencia y su temperatura. Las segundas, las propiedades químicas, producen las sensaciones de frialdad de la menta, lo picoso del chile, etcétera.2
La lengua humana no es sensible de manera uniforme a lo largo de su superficie. Así, la parte media no es sensible al gusto como lo son otras partes. En la figura 60 vemos que la punta de la lengua es sensible principalmente a lo dulce. A los lados y en la parte anterior de la lengua se encuentra la sensibilidad a lo salado. La lengua es principalmente sensible a los sabores agrios a los lados, pero en la parte posterior. Finalmente, la parte posterior central de la lengua es sensible principalmente a los sabores amargos.2


Agrio
Los sabores agrios los percibimos principalmente en sustancias que son ácidas. Estos compuestos contienen átomos de hidrógeno, que son los principales responsables de dicho sabor. Cuando se mezcla la sustancia que contiene el ácido con agua (recuérdese que para que las yemas gustativas reaccionen, las sustancias tienen que estar húmedas), en general desprenden algunos de sus átomos de hidrógeno. Estos átomos de hidrógeno han perdido sus electrones y por lo tanto quedan eléctricamente cargados (Figura 61); se dice que están ionizados. Cuando los átomos de hidrógeno ionizados entran en contacto con las yemas gustativas dan la sensación de un sabor agrio. 1
Sin embargo, este factor no determina exclusivamente el carácter agrio de una sustancia. Por ejemplo, el ácido cítrico que existe en la naranja, el limón, etc., tiene un sabor agrio muy pronunciado que no se puede explicar con base en la cantidad de iones de hidrógeno que produce. Otro ejemplo es el caso del vinagre, que está compuesto de ácido acético y que es mucho más agrio de lo que se esperaría con base en la cantidad de iones de hidrógeno que produce.1
Amargo
En un buen número de casos, las sustancias amargas son sales inorgánicas de alto peso molecular; esto quiere decir que son moléculas que tienen muchos átomos. Otros casos son moléculas orgánicas que tienen añadidas cadenas carbónicas. Por ejemplo, en la figura 62 se muestran tres moléculas que tienen los mismos átomos pero que difieren en su distribución en el espacio; sin embargo, una de ellas no tiene sabor, la segunda es ligeramente amarga y la tercera es dulce. 1
Salado
La sal común o de mesa es un compuesto formado por átomos de sodio y de cloro, el cloruro de sodio. Resulta que el sabor salado que da esta sustancia no se debe exclusivamente ni al sodio ni al cloro. Existen sustancias que contienen cloro y no sodio que son saladas, así como compuestos de sodio que no contienen cloro y que también son salados.1
Los compuestos llamados sales de bajo peso molecular, es decir, con pocos átomos, tienen sabor predominantemente salado y como se vio arriba, las sales con alto peso molecular son principalmente amargas.1
Existen también otras sales que contienen metales. Por ejemplo, sales de mercurio que tienen un sabor metálico mientras que algunas sales de plomo llegan a tener sabor ¡dulce! 1
Dulce
En general, las sustancias que nos dan la impresión de tener sabor dulce están formadas de compuestos orgánicos como los alcoholes, azúcares, glicoles, etc. La sensación de dulce también se debe a la forma en que estén arreglados los átomos en las moléculas; en la figura 62 vimos compuestos con diferentes disposiciones de los mismos átomos, pero que tienen diferentes sabores, uno de ellos dulce.1
Bibliografía
1) http://bibliotecadigital.ilce.edu.mx/sites/ciencia/volumen2/ciencia3/073/htm/sec_10.htm
2) LaTorre, Ramón. 1996. Biofisica y fisiología celular. pp 537.
2) LaTorre, Ramón. 1996. Biofisica y fisiología celular. pp 537.
El SENTIDO DEL OLFATO
OLFATO

Cuando un objeto, emite un olor y nos llega a la nariz esto quiere decir que moléculas de dicho objeto se han desprendido de él y llegado, por difusión o arrastre, a nuestra nariz. Estas moléculas entran en la nariz debido a la aspiración que realizamos cuando respiramos. En este proceso, el aire que inhalamos arrastra a las moléculas que están en la vecindad de la nariz. La corriente de aire que entra da lugar a una corriente secundaria que pasa por el epitelio sensitivo. La cantidad de aire y por tanto la fracción de moléculas del objeto oloroso que se deposita en el epitelio es muy pequeña. 1
También llegan corrientes de aire desde la boca. La comida que tenemos en la boca también despide moléculas que son arrastradas hasta el epitelio sensitivo. Este hecho tiene como consecuencia que la sensación predominante al comer provenga no del gusto que se inicia en la lengua sino del olfato. 1
La sensación de oler se experimenta cuando las moléculas aromáticas llegan a la mucosa nasal, en donde se disuelven. Así, estas moléculas entran en contacto con los cilios.
Hasta hoy en día no se ha podido determinar con certeza el mecanismo por medio del cual se inicia el proceso a través de los receptores nerviosos que nos dan la sensación de oler. Mencionaremos algunas ideas que se han expuesto y que parecen tener algunos elementos que pueden ser verdaderos. 1
En primer lugar, en general, somos muy sensibles a una cantidad extraordinariamente grande de olores distintos. A primera vista podríamos pensar que hay un nervio olfatorio sensible a cada olor posible. Sin embargo, hasta hoy en día no se han encontrado estas diferencias en los cilios nasales. 2
En muchos casos resulta que la presencia de un átomo particular en la molécula de la sustancia olorosa es la que le da su olor peculiar. Por ejemplo, está el caso del agua, cuya molécula tiene un átomo de oxígeno y dos de hidrógeno, H2O. Si se sustituye el átomo de oxígeno por uno de azufre se obtiene el sulfito de hidrógeno, H2S. Esta última sustancia despide el olor de huevos podridos. Vemos que la sustitución de un átomo de oxígeno por uno de azufre lleva a cambios notablemente desagradables en el olor de las sustancias. 3
Por otro lado, existen algunas evidencias de que no es nada más la composición química específica de las moléculas olorosas, sino también su forma la que hace que reaccionemos a su olor. En efecto, existen sustancias cuyas moléculas tienen composiciones químicas diferentes pero con formas muy parecidas, y sentimos que tienen olores muy parecidos.
Sin embargo, la explicación desde el punto de vista molecular de cómo olemos sigue sin respuesta definitiva. 3
La nariz tiene varias funciones primordiales: por una parte constituye el órgano del sentido del olfato, pues el hombre es capaz de discriminar entre dos mil y cuatro mil olores. Además, forma parte de la vía respiratoria filtrando, calentando, humedeciendo el aire y expulsando materiales extraños recogidos por el aire. 1
Por nariz se entiende la pirámide nasal visible en la cara o nariz propiamente tal y su cavidad, la cavidad nasal, que se extiende desde los orificios nasales externos por delante hasta las coanas u orificios nasales posteriores que comunican la cavidad nasal con la nasofaringe. La cavidad nasal está dividida en dos mitades por el tabique nasal; a cada mitad se le conoce como fosa nasal; de su pared externa ósea se originan dos láminas óseas perpendiculares llamadas cornetes superior y medio, que dividen parcialmente a cada fosa en cavidades más pequeñas llamadas meatos: bajo el cornete superior está el meato superior; bajo el cornete medio está el meato medio; una tercera lámina ósea independiente, llamada cornete inferior, origina el meato inferior, cavidad cuyo techo es el cornete inferior, y cuyo piso es el piso de (a fosa nasal; en el meato inferior termina el conducto lacrimonasal, que comunica el ángulo interno del ojo con la nariz. Cada fosa nasal se une por medio de orificios en su pared ósea externa con los llamados senos paranasales, cavidades que se encuentran en el interior de los huesos maxilares, frontal, esfenoides y etmoides. 1
La cavidad nasal presenta tres zonas:
Por nariz se entiende la pirámide nasal visible en la cara o nariz propiamente tal y su cavidad, la cavidad nasal, que se extiende desde los orificios nasales externos por delante hasta las coanas u orificios nasales posteriores que comunican la cavidad nasal con la nasofaringe. La cavidad nasal está dividida en dos mitades por el tabique nasal; a cada mitad se le conoce como fosa nasal; de su pared externa ósea se originan dos láminas óseas perpendiculares llamadas cornetes superior y medio, que dividen parcialmente a cada fosa en cavidades más pequeñas llamadas meatos: bajo el cornete superior está el meato superior; bajo el cornete medio está el meato medio; una tercera lámina ósea independiente, llamada cornete inferior, origina el meato inferior, cavidad cuyo techo es el cornete inferior, y cuyo piso es el piso de (a fosa nasal; en el meato inferior termina el conducto lacrimonasal, que comunica el ángulo interno del ojo con la nariz. Cada fosa nasal se une por medio de orificios en su pared ósea externa con los llamados senos paranasales, cavidades que se encuentran en el interior de los huesos maxilares, frontal, esfenoides y etmoides. 1
La cavidad nasal presenta tres zonas:
1- el vestíbulo: es la parte más anterior e inferior de la nariz, y está tapizada por piel. 2
2- la llamada "región respiratoria": que se continúa con el vestíbulo y se comunica con la nasofaringe, y está tapizada por una membrana mucosa de tipo respiratorio. 2
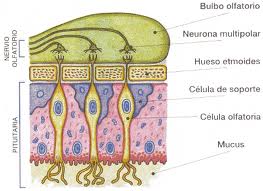
3- la región olfatoria: ubicada en el cornete superior y tercio superior del tabique nasal, donde llegan filetes nerviosos que atraviesan los orificios de un hueso llamado etmoides, cuyo conjunto origina el nervio olfatorio, que alcanza el llamado bulbo olfatorio, que se continúa con las llamadas cintillas olfatorias, cuyas fibras llevan los estímulos al cerebro y cerebelo. 2


{kind=link}
Los receptores químicos del olfato son:
La glándula pituitaria roja: Se ubica en la parte inferior de la fosa nasal y está recubierto por numerosos vasos sanguíneos que calientan el aire. 2
La glándula pituitaria amarilla: Se ubica en la parte superior de las fosas nasales y presenta tres capas:
Células de sostén
Células olfatorias
Células basales
2
Las células olfatorias son células nerviosas receptoras de estímulos químicos provocados por los vapores. En la pituitaria amarilla se encuentran las glándulas mucosas de Bowman, que liberan un líquido que mantiene húmedo y limpio el epitelio olfatorio. 3
Para estimular estas es necesario que las sustancias sean volátiles, es decir, han de desprender vapores que puedan penetrar en las fosas nasales, y que sean solubles en agua para que se disuelvan en el moco y lleguen a las células olfatorias. Éstas transmiten un impulso nervioso al bulbo olfatorio y de este a los centros olfatorios de la corteza cerebral, que es donde se aprecia e interpreta la sensación de olor. 3
Se cree que existen 7 tipos de células olfatorias, cada una de las cuales sólo es capaz de detectar un tipo de moléculas, éstas son:
Alacanforado.
Almizclado.
Floral.
Mentolado.
Etéreo.
Picante.
Pútrido
La glándula pituitaria roja: Se ubica en la parte inferior de la fosa nasal y está recubierto por numerosos vasos sanguíneos que calientan el aire. 2
La glándula pituitaria amarilla: Se ubica en la parte superior de las fosas nasales y presenta tres capas:
Células de sostén
Células olfatorias
Células basales
2
Las células olfatorias son células nerviosas receptoras de estímulos químicos provocados por los vapores. En la pituitaria amarilla se encuentran las glándulas mucosas de Bowman, que liberan un líquido que mantiene húmedo y limpio el epitelio olfatorio. 3
Para estimular estas es necesario que las sustancias sean volátiles, es decir, han de desprender vapores que puedan penetrar en las fosas nasales, y que sean solubles en agua para que se disuelvan en el moco y lleguen a las células olfatorias. Éstas transmiten un impulso nervioso al bulbo olfatorio y de este a los centros olfatorios de la corteza cerebral, que es donde se aprecia e interpreta la sensación de olor. 3
Se cree que existen 7 tipos de células olfatorias, cada una de las cuales sólo es capaz de detectar un tipo de moléculas, éstas son:
Alacanforado.
Almizclado.
Floral.
Mentolado.
Etéreo.
Picante.
Pútrido
3
Transducción olfativa
Las sustancias odorantes son compuestos químicos volátiles transportados por el aire. Los objetos olorosos liberan a la atmósfera pequeñas moléculas que percibimos al inspirar. Estas moléculas alcanzan la mucosa olfativa, que consta de tres tipos característicos de células: las células olfativas sensoriales, las células de sostén y las células basales, que se dividen aproximadamente una vez al mes y reemplazan a las células olfativas moribundas. Los 20 o 30 millones de células olfativas humanas contienen, en su extremo anterior, una pequeña cabeza con cerca de 20 cilios. El moco nasal acuoso transporta las moléculas aromáticas a los cilios con ayuda de proteínas fijadoras; los cilios transforman las señales químicas de los distintos aromas en respuestas eléctricas. 1
Las prolongaciones nerviosas de las células olfativas alcanzan el bulbo olfatorio a través de micro-orificios del cráneo; el bulbo es una porción anterior del cerebro, que se ocupa de la percepción de los olores. Estas prolongaciones nerviosas terminan en los glomérulos, pequeñas terminaciones de células olfativas de forma esférica donde se procesan las señales aromáticas que luego son conducidas por células receptoras especiales. La información llega primero al sistema límbico y al hipotálamo, regiones cerebrales responsables de las emociones, sentimientos, instintos e impulsos, tales regiones almacenan también los contenidos de la memoria y regulan la liberación de hormonas. Por este motivo, los olores pueden modificar directamente nuestro comportamiento y las funciones corporales. Sólo más tarde parte de la información olorosa alcanza la corteza cerebral y se torna consciente. 1
Las sustancias odorantes son compuestos químicos volátiles transportados por el aire. Los objetos olorosos liberan a la atmósfera pequeñas moléculas que percibimos al inspirar. Estas moléculas alcanzan la mucosa olfativa, que consta de tres tipos característicos de células: las células olfativas sensoriales, las células de sostén y las células basales, que se dividen aproximadamente una vez al mes y reemplazan a las células olfativas moribundas. Los 20 o 30 millones de células olfativas humanas contienen, en su extremo anterior, una pequeña cabeza con cerca de 20 cilios. El moco nasal acuoso transporta las moléculas aromáticas a los cilios con ayuda de proteínas fijadoras; los cilios transforman las señales químicas de los distintos aromas en respuestas eléctricas. 1
Las prolongaciones nerviosas de las células olfativas alcanzan el bulbo olfatorio a través de micro-orificios del cráneo; el bulbo es una porción anterior del cerebro, que se ocupa de la percepción de los olores. Estas prolongaciones nerviosas terminan en los glomérulos, pequeñas terminaciones de células olfativas de forma esférica donde se procesan las señales aromáticas que luego son conducidas por células receptoras especiales. La información llega primero al sistema límbico y al hipotálamo, regiones cerebrales responsables de las emociones, sentimientos, instintos e impulsos, tales regiones almacenan también los contenidos de la memoria y regulan la liberación de hormonas. Por este motivo, los olores pueden modificar directamente nuestro comportamiento y las funciones corporales. Sólo más tarde parte de la información olorosa alcanza la corteza cerebral y se torna consciente. 1

Hasta hoy en día no se ha podido determinar con certeza el mecanismo por medio del cual se inicia el proceso a través de los receptores nerviosos que nos dan la sensación de oler. Mencionaremos algunas ideas que se han expuesto y que parecen tener algunos elementos que pueden ser verdaderos. 2
En primer lugar, en general, somos muy sensibles a una cantidad extraordinariamente grande de olores distintos. A primera vista podríamos pensar que hay un nervio olfatorio sensible a cada olor posible. Sin embargo, hasta hoy en día no se han encontrado estas diferencias en los cilios nasales. 2
En muchos casos resulta que la presencia de un átomo particular en la molécula de la sustancia olorosa es la que le da su olor peculiar. Como ilustración podemos mencionar el caso del agua, cuya molécula tiene un átomo de oxígeno y dos de hidrógeno, H2O. Si se sustituye el átomo de oxígeno por uno de azufre se obtiene el sulfito de hidrógeno, H2S. Esta última sustancia despide el olor de huevos podridos. Otro ejemplo es el del bióxido de carbono que tiene un átomo de carbón y dos de oxígeno. Su fórmula química es CO2. Este gas no tiene olor. Si ahora reemplazamos los oxígenos por átomos de azufre obtenemos el disulfuto de carbono, cuya fórmula química es CS2. Resulta que el olor de este último compuesto nos es repelente. Vemos que la sustitución de un átomo de oxígeno por uno de azufre lleva a cambios notablemente desagradables en el olor de las sustancias. 2
En primer lugar, en general, somos muy sensibles a una cantidad extraordinariamente grande de olores distintos. A primera vista podríamos pensar que hay un nervio olfatorio sensible a cada olor posible. Sin embargo, hasta hoy en día no se han encontrado estas diferencias en los cilios nasales. 2
En muchos casos resulta que la presencia de un átomo particular en la molécula de la sustancia olorosa es la que le da su olor peculiar. Como ilustración podemos mencionar el caso del agua, cuya molécula tiene un átomo de oxígeno y dos de hidrógeno, H2O. Si se sustituye el átomo de oxígeno por uno de azufre se obtiene el sulfito de hidrógeno, H2S. Esta última sustancia despide el olor de huevos podridos. Otro ejemplo es el del bióxido de carbono que tiene un átomo de carbón y dos de oxígeno. Su fórmula química es CO2. Este gas no tiene olor. Si ahora reemplazamos los oxígenos por átomos de azufre obtenemos el disulfuto de carbono, cuya fórmula química es CS2. Resulta que el olor de este último compuesto nos es repelente. Vemos que la sustitución de un átomo de oxígeno por uno de azufre lleva a cambios notablemente desagradables en el olor de las sustancias. 2
Bibliografía
1) Fox, Stuart Ira.2003. Fisiología humana. pp. 254-255
2) http://www.mancia.org/foro/fisiologia-biofisica/54859-fisiologia-sensorial.html
3) http://www.hhmi.org/news/zuker4-esp.html
2) http://www.mancia.org/foro/fisiologia-biofisica/54859-fisiologia-sensorial.html
3) http://www.hhmi.org/news/zuker4-esp.html
domingo, 17 de julio de 2011
Audición
Sistema Auditivo

La audición se desarrolla en tres etapas básicas:
• Captación y procesamiento mecánico. 2
• Conversión de la señal acústica en impulsos nerviosos. 2
• Procesamiento neural.2
La captación, procesamiento y transducción de los estímulos sonoros se lleva acabo en el oído, mientras que la etapa del procesamiento neural, en el que se producen las sensaciones auditivas, se encuentra ubicada en el cerebro. 2
Así, se distinguen dos regiones del sistema audito: La región periférica y la región central. 2
Región Periférica
El oído o región periférica se divide en tres zonas: oído externo, oído medio y oído interno. 2

Oído externo
Su función es recolectar las ondas sonoras y encauzarlas al oído medio. 2
Pabellón auricular u oreja: Dirige las ondas sonoras hacia el conducto auditivo externo a través del orificio auditivo. 2
Conducto auditivo externo: Tubo de 2cm de longitud que protege las delicadas estructuras del oído medio y minimiza la distancia del oído interno al cerebro, reduciendo el tiempo de propagación de los impulsos nerviosos. 2

Las señales sonoras que entran al conducto auditivo externo sufren efectos de difracción y estos varían de acuerdo a la dirección de incidencia. Esto determina la procedencia del sonido. 2
Oído medio
Constituido por una cavidad llena de aire, donde se localizan 3 huesecillos: Martillo, yunque y estribo unidos de forma articulada. Uno de los extremos del martillo se encuentra unido a la membrana timpánica, mientras el estribo se une a la ventana oval a través de un anillo flexible. Orificio que constituye la vía de entrada del sonido al oído interno. 2
La cavidad del oído medio se comunica con la trompa de Eustaquio, la cual es un conducto que llega hasta las vías respiratorias y permite igualar la presión de aire a ambos lados del la membrana timpánica. 2


Oído interno
Es el final de la cadena de procesamiento mecánico del sonido. 2
Se lleva a cabo:
• Filtraje de la señal sonora.
• Transducción.
• Generación de impulsos.
2
En el oído interno se encuentra:
La coclea o caracol, que consta de:
• Conducto rígido en forma de espiral
• Aproximadamente 35 mm de longitud
• Dividido longitudinalmente por la membrana basilar y la membrana de Reissner.
• Consta de 3 compartimientos o escalas: Escala vestibular y la escala timpánica que contienen el mismo fluido (perilinfa), ya que se conectan por una pequeña abertura en el vértice (helicotrema). La escala media que contiene como fluido endolinfa.
2

La base del estribo, a través de la ventana oval, está en contacto con el fluido de la escala vestibular, mientras que la escala timpánica desemboca en la cavidad del oído medio a través de otra abertura (ventana redonda) sellada por una membrana flexible (membrana timpánica secundaria). 2

Órgano de Corti
Localizado en la rampa coclear o media del oído interno de los mamíferos y compuesto por las células sensoriales auditivas llamadas células ciliadas (transductores de señales sonoras a impulsos nerviosos). 2
Esta entre la membrana basilar y la membrana tectorial. 2


Se pueden distinguir dos tipos de células ciliares: Externas e internas. 2
Ambos tipos presentan conexiones y sinapsis con las fibras nerviosas aferentes y eferentes, las cuales conforman el nervio auditivo. 2
• 90% de las fibras aferentes inervan a las células ciliares internas. 2
• La mayoría de fibras eferentes inervan a las células ciliares externas. 2

Propagación del sonido en la cóclea
La membrana de Reissner, la cual separa los fluidos de la escala vestibular y media, es sumamente delgada y en concecuencia, los líquidos de ambas escalas pueden tratarse como uno solo desde el punto de vista de la dinámica de fluidos. 2
Así, las oscilaciones de la perilinfa de la escala vestibular se trasmiten a la endolinfa y esta a la membrana basilar que a su vez provoca oscilaciones en el fluido de la escala timpánica. 2
La propagación de las oscilaciones del fluido en la escala vestibular a la timpánica no sólo se lleva a cabo a través de la membrana basilar; para sonidos de muy baja frecuencia, las vibraciones se transmiten a través del helicotrema. 2

En conclusión, el sonido propagado a través del oído externo y medio llega hasta la cóclea, donde las oscilaciones en los fluidos hacen vibrar a la membrana basilar y a todas las estructuras que ésta soporta. 2
La cóclea como analizador de frecuencias
La membrana basilar es una estructura cuyo espesor y rigidez no es constante: cerca de la ventana oval, la membrana es gruesa y rígida, pero a medida que se acerca hacia el vértice de la cóclea se vuelve más delgada y flexible. 2
La rigidez decae casi exponencialmente con la distancia a la ventana oval; esta variación de la rigidez en función de la posición afecta la velocidad de propagación de las ondas sonoras a lo largo de ella, y es responsable en gran medida de un fenómeno muy importante: la selectividad en frecuencia del oído interno. 2

Mecanismo de transducción
El proceso de transducción o conversión de señal mecánica a electroquímica se desarrolla en el órgano de Corti, situado sobre la membrana basilar. 2
Las vibraciones de la membrana basilar hacen que ésta se mueva en sentido vertical. A su vez la membrana tectorial, ubicada sobre las células ciliares vibra igualmente; sin embargo, dado que los ejes de movimiento de ambas membranas son distintos, el efecto final es el de un desplazamiento "lateral" de la membrana tectorial con respecto a la membrana basilar. 2
Como resultado, los cilios de las células ciliares externas se "doblan" hacia un lado u otro . 2
En el caso de las células internas, aun cuando sus cilios no están en contacto directo con la membrana tectorial, los desplazamientos del líquido y su alta viscosidad hacen que dichos cilios se doblen también. 2
El proceso de transducción en las células pilosas se ha estudiado con técnicas electrofisiológicas y otras que han permitido entender el mecanismo iónico involucrado. 2
En la punta de los cilios de las células pilosas existen canales de K+, catión que es muy abundante en la endolinfa, líquido que esta en contacto con dichas células. En condiciones de reposo ellas presentan un potencial de membrana que fluctúa entre 452 y 60
mV, con respecto a la endolinfa. Esos canales de K+ están abiertos en bajo número lo que explicaría la variabilidad del potencial de reposo ya que estaría entrando ese catión y tendiendo a despolarizar a la célula.
Los cilios se mueven debido a la influencia de las ondas que vienen por la perilinfa. 2
Cuando se mueven se abren más canales de transducción lo que provoca un mayor entrada de K+, con la consiguiente despolarización de las células. Esta disminución de su potencial de reposo abre canales de Ca2+dependientes de voltaje lo cual gatilla la liberación del neurotransmisor que excita a un grupo de terminales nerviosos que inervan dichas células. Estos responden generando potenciales de acción que viajan por la vía auditiva hasta el sistema nervioso central. 1

Las fibras nerviosas aferentes llevan información a diversos lugares del cerebro. En esté se encuentran estructuras de mayor o menos complejidad, encargadas de procesar distintos aspectos de la información
Por ejemplo, en los centros “inferiores” del cerebro se recibe, procesa e intercambia información proveniente de ambos oídos, con el fin de determinar la localización de las fuentes de sonido. En los centros “superiores” de la corteza existen estructuras más especializadas que responden a estímulos más complejos. 1
La información transmitida por el nervio auditivo se utiliza finalmente para generar lo que se conoce como “sensaciones”.
1
Suscribirse a:
Entradas (Atom)